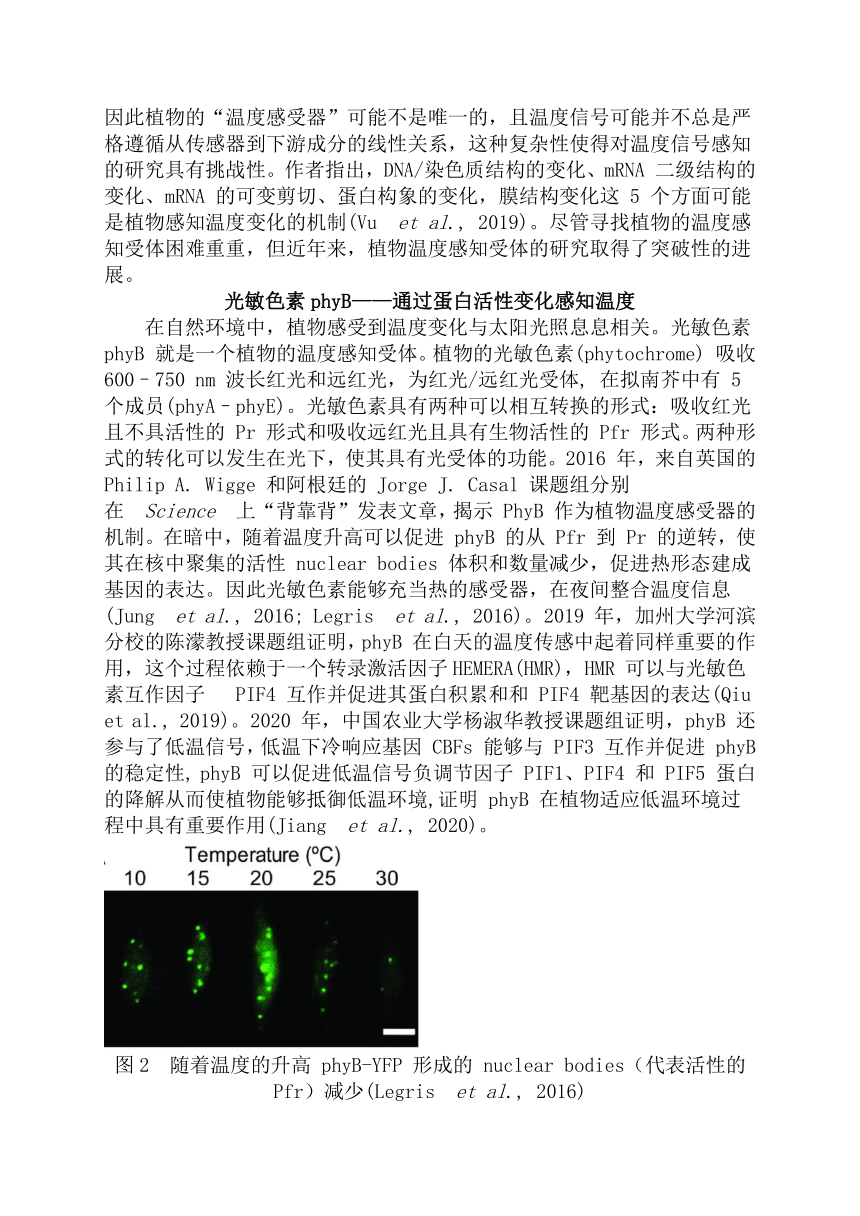
資源簡介 植物對溫度的感知2019版高中生物學選擇性必修一說,溫度參與調節植物的生長發育:那么,植物是如何感知溫度的呢?地球較為合適的氣候為生命提供了繁衍生息的最佳場所,地球的氣溫會隨著太陽輻射、緯度、海陸位置等因素而產生變化。在生物進化的過程中,動、植物分別進化出了不同的機制來感知溫度的變化。動物細胞主要通過膜上的 TRP 家族離子通道來作為溫度的感知受體。1997 年 David Julius 教授成功篩選并克隆到辣椒素受體 VR1(TRPV1),并證明 TRPV1 不僅能夠被辣椒素特異激活,還能被 42 °C 以上高溫激活。隨后一系列 TRP家族成員被鑒定為溫度感受器(ThermoTRPs),這些成員在一定的溫度下被激活,共同感知整個生理范圍內的溫度,其中一些家族成員還可以被薄荷醇、大蒜素等化學配體結合并激活(Castillo et al., 2018)。圖1 感知不同溫度的 TRPs 通道(Castillo et al., 2018)然而,植物基因組中未發現編碼 TRP 家族基因,暗示在植物中存在不同的蛋白或不同于動物細胞的溫度感知機制。2019 年 Lam Dai Vu 等人的觀點性綜述指出,理論上溫度可以影響所有生化過程的吉布斯自由能平衡,因此植物的“溫度感受器”可能不是唯一的,且溫度信號可能并不總是嚴格遵循從傳感器到下游成分的線性關系,這種復雜性使得對溫度信號感知的研究具有挑戰性。作者指出,DNA/染色質結構的變化、mRNA 二級結構的變化、mRNA 的可變剪切、蛋白構象的變化,膜結構變化這 5 個方面可能是植物感知溫度變化的機制(Vu et al., 2019)。盡管尋找植物的溫度感知受體困難重重,但近年來,植物溫度感知受體的研究取得了突破性的進展。光敏色素phyB——通過蛋白活性變化感知溫度在自然環境中,植物感受到溫度變化與太陽光照息息相關。光敏色素 phyB 就是一個植物的溫度感知受體。植物的光敏色素(phytochrome) 吸收 600–750 nm 波長紅光和遠紅光,為紅光/遠紅光受體, 在擬南芥中有 5 個成員(phyA–phyE)。光敏色素具有兩種可以相互轉換的形式:吸收紅光且不具活性的 Pr 形式和吸收遠紅光且具有生物活性的 Pfr 形式。兩種形式的轉化可以發生在光下,使其具有光受體的功能。2016 年,來自英國的 Philip A. Wigge 和阿根廷的 Jorge J. Casal 課題組分別在 Science 上“背靠背”發表文章,揭示 PhyB 作為植物溫度感受器的機制。在暗中,隨著溫度升高可以促進 phyB 的從 Pfr 到 Pr 的逆轉,使其在核中聚集的活性 nuclear bodies 體積和數量減少,促進熱形態建成基因的表達。因此光敏色素能夠充當熱的感受器,在夜間整合溫度信息(Jung et al., 2016; Legris et al., 2016)。2019 年,加州大學河濱分校的陳濛教授課題組證明,phyB 在白天的溫度傳感中起著同樣重要的作用,這個過程依賴于一個轉錄激活因子HEMERA(HMR),HMR 可以與光敏色素互作因子 PIF4 互作并促進其蛋白積累和和 PIF4 靶基因的表達(Qiu et al., 2019)。2020 年,中國農業大學楊淑華教授課題組證明,phyB 還參與了低溫信號,低溫下冷響應基因 CBFs 能夠與 PIF3 互作并促進 phyB 的穩定性, phyB 可以促進低溫信號負調節因子 PIF1、PIF4 和 PIF5 蛋白的降解從而使植物能夠抵御低溫環境,證明 phyB 在植物適應低溫環境過程中具有重要作用(Jiang et al., 2020)。圖2 隨著溫度的升高 phyB-YFP 形成的 nuclear bodies(代表活性的 Pfr)減少(Legris et al., 2016)ELF3——通過相分離感知高溫溫度變化是植物生物鐘感知的一部分。擬南芥生物鐘晚間轉錄抑制復合物(evening complex) 不僅參與了生物鐘的調控,還參與了溫度響應。這一復合物包含 ELF3、ELF4 和 LUX 三個蛋白,可以在晚間抑制 PIF4 基因的表達。高溫能夠解除復合物對于 PIF4 基因的抑制,然而其機制尚不清楚。2019 年英國劍橋大學 Philip A. Wigge 課題組在 Nature 發表文章揭示高溫通過促進 ELF3 的相分離調控植物的熱響應。ELF3 含有多聚谷氨酰胺的重復序列(polyQ),為預測的朊病毒類似結構域(PrD),polyQ 的長度影響 ELF3 的活性。進一步發現,ELF3 在高溫下會聚集形成點狀,并且這種聚集的形成依賴于 polyQ,隨著 polyQ 長度的增加形成亮點的趨勢越大。體外實驗進一步證明,ELF3 的 PrD 結構域可以隨著溫度升高可逆地形成液滴,說明溫度可以促進 ELF3 的相分離。本文揭示了一種新的熱感性機制:溫度通過調控 ELF3 的相變在活動和非活動狀態之間快速轉變。有意思的是,各種擬南芥的生態型和生活在不同地方的植物具有不同長度 polyQ 結構域,這與不同植物的耐熱性相關。(Jung et al., 2020)圖3 植物通過 ELF3 的相分離響應高溫促使擬南芥開花(Alberti, 2020)COLD1——植物低溫感受器2015年中科院植物研究所種康院士課題組報道發現水稻 COLD1 為植物低溫感受器。COLD1 編碼一種定位于質膜和內質網的跨膜蛋白,其突變導致水稻對冷害敏感。研究發現,COLD1 能與植物中 G 蛋白異源三聚體的 α 亞基RGA1 發生互作,促進 G 蛋白 GTP 酶活性,并激活響應低溫的 Ca2+ 通道。COLD1 基因第四個外顯子的單個核苷酸的突變影響秈稻、粳稻的耐寒性差異,同時還揭示了遺傳進化過程中,來源于中國野生稻品種的 O. refipogon的 CLOD1 基因賦予粳稻的耐寒性(Ma et al., 2015)。近期,種康院士課題組進一步揭示 COLD1 通過調控下游維生素 E-維生素 K1 的代謝調控水稻的低溫耐受性。然而目前對 COLD1 感知低溫的分子機制及受 COLD1 激活的 Ca2+ 通道仍不清楚(Luo et al., 2021)。未來,需要通過結構生物學等手段進一步闡釋低溫激活 COLD1 的原理和構象變化。圖4 粳稻在馴化過程中通過 COLD1 基因位點的選擇來適應低溫(Li and Lin, 2016)植物固著生長的特點迫使植物進化出更加復雜的溫度感應機制以應對復雜的環境變化。對植物溫度感知的研究將有助于我們認識植物適應環境的機制,為作物品種改良提供重要的理論基礎。例、下列關于植物感溫性的描述,正確的是( )A.感溫性是植物對環境溫度規律性變化的適應B.在一定溫度范圍內,隨溫度升高,植物生長發育速度加快C.有些植物或品種在較高溫度刺激下發育加快,即感溫性較弱D.晚稻的感溫性強于中稻,中稻強于早稻答案:ABD 展開更多...... 收起↑ 資源預覽 縮略圖、資源來源于二一教育資源庫
 資源預覽
資源預覽

 資源預覽
資源預覽